Kimberella quadrata Mikhail Fedonkin e Ben Waggoner
Phylum: Mollusca Linnaeus, 1758
Subphylum:
Classe:
Ordine: Bilateria Hatschek , 1888
Famiglia:
Genere: Kimberella Wade, 1972
Specie e sottospecie
Il genere annovera attualmente le seguenti specie: Kimberella persii Vaziri, Majidifard & Laflamme, 2018 - Kimberella quadrata Glaessner & Wade, 1966.
Descrizione
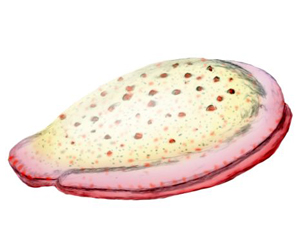
Il genere è stato chiamato così in onore del signor John Kimber, studente, insegnante e collezionista; che perse la vita durante una spedizione nell'Australia centrale nel 1964. In origine, al genere fu dato il nome Kimberia, tuttavia, il nome Kimberia era già in uso come sottogenere di Turritella (Gastropoda), secondo il dottor NH Ludbrook; e un nuovo genere, Kimberella, fu proposto da Mary Wade nel 1972. Oltre 1.000 esemplari, rappresentanti organismi di tutti gli stadi di maturità, sono stati ora trovati nell'area del Mar Bianco sul fondo di strati di arenaria a grana fine. Il gran numero di esemplari, la piccola granulometria dei sedimenti e la varietà di circostanze in cui gli esemplari sono stati conservati forniscono informazioni dettagliate sulla forma esterna, l'anatomia interna, la locomozione e lo stile di alimentazione della Kimberella. Tutti i fossili hanno un contorno ovale. Gli esemplari allungati illustrano che l'organismo era in grado di allungarsi in direzione antero-posteriore, forse di un fattore due. Come molti altri esemplari trovati nel Mar Bianco, il tipo più comune di simmetria osservato sembra essere bilaterale; con poco o nessun segno di alcun tipo di simmetria radiale trovata nei Cnidari, il gruppo che include meduse, anemoni di mare e idre. I fossili australiani erano originariamente descritti come un tipo di medusa, ma questo è incoerente con la simmetria bilaterale osservata nei fossili. I fossili del Mar Bianco e i sedimenti circostanti mostrano anche che Kimberella viveva sulla superficie del fondale marino. La Kimberella aveva una copertura dorsale che è stata descritta come un "guscio morbido" (non mineralizzato); negli esemplari più grandi, questo raggiungeva fino a 15 cm di lunghezza, 5-7 cm di larghezza e 3-4 cm di altezza; con uno spessore minimo di 2-3 mm. Il guscio era rigido ma flessibile e sembra non essere stato mineralizzato, diventando più duro man mano che diventava più grande (e presumibilmente più spesso) negli esemplari più maturi. La deformazione osservata negli esemplari allungati e piegati illustra che il guscio era altamente malleabile; forse, piuttosto che un singolo tegumento, consisteva in un'aggregazione di scleriti mineralizzati. Nel punto più alto c'era una struttura simile a un cappuccio, che formava quella che si pensa fosse la parte anteriore. In alcuni esemplari, la superficie interna del guscio presenta strisce che si estendono per tutta la larghezza della creatura; queste possono rappresentare punti di attacco muscolare. Strisce simili attorno al bordo del guscio potrebbero essere state collegate ai muscoli che ritraevano il piede nel guscio. L'asse lungo dell'organismo è segnato da una cresta rialzata; l'asse centrale è leggermente gibboso. Il corpo della Kimberella non aveva alcuna segmentazione visibile ma aveva una serie di " moduli " ripetuti. Ogni modulo includeva una fascia ben sviluppata di muscoli dorso-ventrali che andavano dalla parte superiore al singolo, largo e muscoloso "piede", e muscoli ventrali trasversali più piccoli lateralmente attraverso la parte inferiore del corpo. La combinazione di queste fasce muscolari dorso-ventrali e ventrali trasversali consentiva alla Kimberella di muoversi increspando il piede. Il corpo aveva anche una frangia arricciata che potrebbe essere stata parte del sistema respiratorio dell'animale, svolgendo una funzione simile a quella delle branchie . Il fatto che la frangia si estendesse ben oltre il guscio potrebbe indicare che le "branchie" della Kimberella erano inefficienti e necessitavano di un'ampia area, o che non c'erano predatori efficaci sulla Kimberella e la funzione principale del guscio era quella di fornire una piattaforma per i muscoli. La Kimberella probabilmente brucava su tappeti microbici, ma non si può escludere un'abitudine predatoria selettiva. Il contenuto intestinale fossilizzato sembra confermare che brucava batteri bentici e alghe, queste ultime avendo attraversato il loro "big bang" ecologico 650 milioni di anni fa nell'ecosistema marino, fornendo una dieta più ricca di nutrienti per i primi animali. Fedonkin ritiene che mentre mangiava, si muoveva "indietro"; la pista così creata veniva distrutta dalla successiva attività di pascolo. Al contrario, Gehling et al. affermano che si muoveva "in avanti". Spesso si trovano ventagli di scanalature che si irradiano dall'estremità della "testa" dell'organismo; queste indicano che l'organismo è rimasto in un posto e ha rastrellato la superficie del tappeto microbico verso di sé tramite l'estensione della sua testa, che portava due "denti". Gehling et al. ricostruirono Kimberella come avente un lungo collo che funzionava come il braccio di un escavatore, ruotando attorno a un asse perpendicolare al fondale marino per produrre la spazzata del ventaglio, e ruotando verso e lontano dall'animale per raschiare il cibo dal substrato alla bocca. In una comunità è stato dimostrato che la Kimberella evitava le sue tracce di pascolo, dimostrando un comportamento sensoriale complesso. La mancanza di prove di riproduzione asessuata suggerisce che gli organismi si siano riprodotti sessualmente. La gemmazione o la fissione non sono mai state osservate.
Diffusione
La Kimberella è stata trovata nelle colline di Ediacara nell'Australia meridionale, nella formazione di Ust' Pinega nella regione del Mar Bianco nella Russia nord-occidentale, nel nord-est del Brasile e nella serie Kushk dell'Iran centrale. I fossili del Mar Bianco sono spesso associati agli "animali" di Ediacara Tribrachidium e Dickinsonia, tracce fossili tortuose, probabilmente prodotte dalla Kimberella e alghe. Gli strati nella successione del Mar Bianco sono stati datati a 555,3 ± 0,3 milioni di anni fa e 558 milioni di anni fa tramite datazione radiometrica, utilizzando rapporti uranio-piombo negli zirconi trovati negli strati di cenere vulcanica che sono schiacciati tra strati che contengono fossili di Kimberella. I fossili di Kimberella sono anche noti da strati sia più vecchi che più giovani di questo intervallo di datazione precisa. I fossili delle colline di Ediacara non sono stati datati con precisione. La Kimberella viveva in acque poco profonde (fino a decine di m di profondità), condividendo il fondale marino calmo e ben ossigenato con organismi fotosintetici e tappeti microbici. Gli assemblaggi che ospitano la Kimberella spesso contengono anche fossili di Andiva, Yorgia, Dickinsonia, Tribrachidium e Charniodiscus, il che suggerisce che vivesse accanto a questi organismi. Le acque in cui viveva la Kimberella erano occasionalmente disturbate da correnti sabbiose , causate quando i sedimenti venivano sollevati dalle tempeste o dallo scarico di acqua di fusione e travolgevano le creature. In risposta a questo stress, gli organismi sembrano aver ritirato le loro parti molli nei loro gusci; apparentemente non potevano muoversi abbastanza velocemente da superare le correnti. Alcuni organismi sopravvissero alla corrente e tentarono di scavare fuori dalla sabbia che era stata depositata sopra di loro; alcuni tentativi infruttuosi possono essere visti dove i giovani erano fossilizzati alla fine di una tana lunga pochi cm.
Bibliografia
Wade, M. (1972). "Hydrozoa and Scyphozoa and other medusoids from the Precambrian Ediacara fauna, South Australia" (PDF). Palaeontology. 15: 197-225.
Fedonkin, M.A.; Simonetta, A; Ivantsov, A.Y. (2007), "New data on Kimberella, the Vendian mollusc-like organism (White sea region, Russia): palaeoecological and evolutionary implications", in Vickers-Rich, Patricia; Komarower, Patricia (eds.), The Rise and Fall of the Ediacaran Biota, Special publications, vol. 286, London: Geological Society, pp. 157-179.
Nelson R Cabej (2019). Epigenetic Mechanisms of the Cambrian Explosion. Elsevier Science. p. 152.
Butterfield, N.J. (2006). "Hooking some stem-group "worms": fossil lophotrochozoans in the Burgess Shale". BioEssays. 28 (12): 1161-6.
Glaessner, M.F.; Wade, M. (1966). "The late Precambrian fossils from Ediacara, South Australia" (Free full text). Palaeontology. 9 (4): 599.
Glaessner, M.; Daily, B. (1959). "The Geology and Late Precambrian Fauna of the Ediacara Fossil Reserve" (PDF). Records of the South Australian Museum. 13: 369-401.
Vaziri, Seyed Hamid; Majidifard, Mahmoud Reza; Laflamme, Marc (2018-03-22). "Diverse Assemblage of Ediacaran fossils from Central Iran". Scientific Reports. 8 (1): 5060.
Martin, M.W.; Grazhdankin, D.V.; Bowring, S.A.; Evans, D.A.D.; Fedonkin, M.A.; Kirschvink, J.L. (2000-05-05). "Age of Neoproterozoic Bilaterian Body and Trace Fossils, White Sea, Russia: Implications for Metazoan Evolution". Science (abstract). 288 (5467): 841-5.
Fedonkin, M. A.; Waggoner, B. M. (1997). "The Late Precambrian fossil Kimberella is a mollusc-like bilaterian organism". Nature. 388 (6645): 868-871.
Jenkins, R.J.F. (1992). "Functional and ecological aspects of Ediacaran assemblages". In Lipps, J.; Signor, P.W. (eds.). Origin and Early Evolution of the Metazoa. New York, NY: Springer. pp. 131-176.
Ivantsov, A.Y. (2009). "A new reconstruction of Kimberella, a problematic Vendian Metazoan". Paleontological Journal. 43 (6): 601-611.
Fedonkin, M.A.; Simonetta, A.; Ivantsov, A.Y. (2007). "New data on Kimberella, the Vendian mollusc-like organism (White Sea region, Russia): Palaeoecological and evolutionary implications" (PDF). Geological Society, London, Special Publications. Special Publications. 286 (1). London, UK: Geological Society: 157-179.
World's oldest meal helps unravel mystery of our earliest animal ancestors.
"The algae that terraformed Earth". BBC News.
Gehling, J. G.; Runnegar, B. N.; Droser, M. L. (2014). "Scratch Traces of Large Ediacara Bilaterian Animals". Journal of Paleontology. 88 (2): 284-298.
Mitchell, E. G.; Bobkov, N.; Dhungana, A.; Kolesnikov, A. V.; Hogarth, I. R. P.; Liu, A. G.; Mustill, T. M. R.; Sozonov, N.; Rogov, S.; Xiao, V. I.; Grazhdankin, D. V. (2020). "The influence of environmental setting on the community ecology of Ediacaran organisms". Interface Focus. 10 (4).
Getty, P.R. (2006). "Producing And Preserving Climactichnites". Geological Society of America Abstracts with Programs. 38 (7): 475.
Andrey Yu. Ivantsov (2013). "Trace Fossils of Precambrian Metazoans "Vendobionta" and "Mollusks"". Stratigraphy and Geological Correlation. 21 (3): 252-264.
Erwin, Douglas H.; Eric H. Davidson (1 July 2002). "The last common bilaterian ancestor". Development. 129 (13): 3021-3032.
Andrey Yu. Ivantsov (2010). "Paleontological evidence for the supposed precambrian occurrence of mollusks". Paleontological Journal. 40 (12): 1552-1559.
Ivantsov, A. Y. (2013). "Trace fossils of precambrian metazoans "Vendobionta" and "Mollusks"". Stratigraphy and Geological Correlation. 21 (3): 252-264.
Smith, M. R. (2012). "Mouthparts of the Burgess Shale fossils Odontogriphus and Wiwaxia: Implications for the ancestral molluscan radula". Proceedings of the Royal Society B. 279 (1745): 4287-4295.
Butterfield, N. J. (2008). "An Early Cambrian Radula". Journal of Paleontology. 82 (3): 543-554.
Cowen, R. (2000). History of Life (3rd ed.). Blackwell Science. p. 63.
Darwin, C (1859). On the Origin of Species by Natural Selection. Murray, London, United Kingdom. pp. 315-316.

|
Data: 21/04/2005
Emissione: Animali preistorici Stato: Macedonia Nota: Emesso in un foglietto di 6 v. diversi |
|---|